 Stato 1994; Roberto Canullo.
Stato 1994; Roberto Canullo.
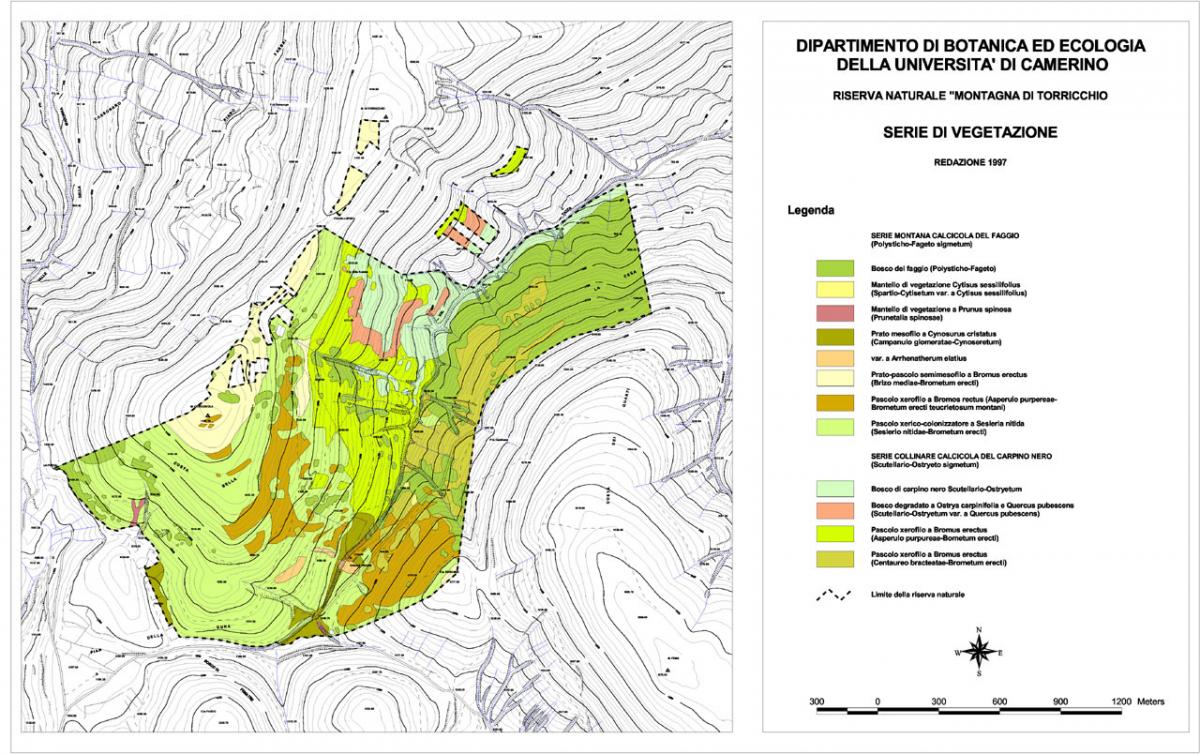
La struttura dei prati pascoli a Cynosurus cristatus
La presente ricerca si riferisce ad un controllo della composizione floristica e della fitomassa specifica dei prati falciabili a Cynosurus cristatus della Riserva Naturale di Torricchio prima del rispristino, avvenuto di recente, delle concimazioni naturali.
Si sono installate due serie di 5 aree ciascuna all’ interno delle quali tutta la vegetazione è stata tagliata a 2 cm dal suolo per simulare lo sfalcio; le dimensioni di ogni quadrato (0,5x0,5) sono conformi agli standard sperimentali generalmente utilizzati per le ricerche sulla vegetazione prativa.
La copertura della vegetazione stimata entro ciascuna delle aree di studio è compresa tra il 95 ed il 100%.
Risultano sempre presenti alcune specie caratteristiche delle unità fitosociologiche superiori (Festuca rubra, Plantago lanceolata, Trifolium pratense, Tragopogon pratensis) in genere con un elevato numero di getti fertili, mentre Cynosurus cristatus manca in una sub-area come pure Bromus hordeaceus, Brachypodium rupestre e Bellis perennis (tutte con pochissimi getti fertili); le altre specie che ottengono il 100% di frequenza sono tutte terofite che non partecipano alla composizione specifica caratteristica dei prati umidi ma che, essendo opportuniste capaci di occupare rapidamente anche i più piccoli spazi effimeri che si aprono nella vegetazione, testimoniano la frequente presenza di "microgaps".
In particolare le componenti fitosociologiche della fitocenosi in questione sono:
Cynosurion e sintaxa superiori 23,4%
Festuco-Brometea 10,6%
Penetrazioni da altri sintaxa
(Agropyretea, Chenopodietea,
Artemisietea, Secalinetea, Sedo-Scleranthetea) 14,9%
Nello spettro biologico le emicriptofite dominano grandemente raggiungendo nel complesso il 68,08% con prevalenza delle emicriptofite scapose e cespitose ed una notevole percentuale di emicriptofite a rosetta (8,50%).
La caratterizzazione ecologica indiretta ha permesso di individuare tre gruppi di specie tipiche di suoli ricchi, mediamente ricchi e poveri a riguardo dei nutrienti (specialmente azoto), rispetto a differenti tenori di umidità del suolo .
Tutti i dati indicano un generale impoverimento della fitocenosi rispetto alle caratteristiche che ci si poteva attendere nei prati a Cynosurus cristatus della Riserva naturale di Torricchio, fatto confermato da analisi fitosociologiche .
La penetrazione sempre più massiccia di specie trasgressive provenienti dai vicini pascoli xerici, il ridotto ruolo (in termini di frequenza, sforzo riproduttivo e biomassa) di Cynosurus cristatus e di altre specie tipiche di queste fitocenosi, assieme alla loro scarsa vitalità, documentano quanto la concessione del solo sfalcio sia stata insufficiente a mantenere la diversità specifica legata ai prati pingui della Riserva.
Appare pertanto giustificato l’ormai consolidato ripristino della concimazione organica che, nelle condizioni di relativa umidità che si verificano nelle aree oggetto di studio, potrà col tempo tornare a favorire le specie che di tali condizioni ecologiche beneficiano. Non appare invece possibile consentire il pascolo estivo-autunnale che normalmente seguiva lo sfalcio, per gli evidenti problemi che ciò comporterebbe nella gestione della Riserva.
La struttura dei pascoli a Sesleria nitida
Stato 1994; Doina Ivan, Nicolae Donita, Dan Gafta, Roberto Canullo.
Sesleria nitida Ten. è una specie endemica dei monti calcarei nel centro e nel sud della penisola e nella Sicilia, distribuita in una fascia altitudinale che va dai 600 ai 2000 m.
Nella Carta della vegetazione della Riserva naturale di Torricchio i prati a sesleria, sia chiusi che aperti, compaiono nell’unità cartografica attribuita ai prati edificati da Festuca ovina. Tali formazioni occupano stazioni esposte ai venti sotto forma di pascoli degradati e aperti e sono state attribuite all’associazione Seslerio nitidae-Brometum erecti, ma senza presentare rilievi.
Il presente contributo si propone di comparare la struttura di due fitocenosi a sesleria, caratterizzate da una diversa copertura (cotica aperta o chiusa) ed esposizione (est-ovest) e stessa pendenza (35°-40°).
Le due fitocenosi messe a confronto si trovano una sul versante ovest del Monte Rotondo fra 1125 e 1175 m di quota (fitocenosi 1), e l’altra sul versante est del Monte Cetrognola fra 1200 e 1250 m di altitudine (fitocenosi 2).
L’origine di tali pascoli è difficile da precisare, ma si possono formulare due ipotesi alternative:
a) formati direttamente in seguito ai disboscamenti su suoli già superficiali;
b) risultato di una regressione dei prati secondari derivati dal disboscamento, dovuta al sovrapascolamento ed alla erosione successiva.
L’analisi della struttura di questi pascoli ha messo in evidenza la loro grande complessità fitocenotica, ma anche l’elevata variabilità delle componenti dal punto di vista del numero di individui, della frequenza, della fitomassa e della potenzialità generativa. Queste caratteristiche sono tipiche per prati sviluppati su suoli erosi, che si trovano in un processo continuo di evoluzione dal prato aperto verso quello chiuso (a cui sembra contribuire anche la colonizzazione arbustiva del citiso) con la contemporanea formazione di nuovo suolo, dovuta anche a processi geomorfologici particolari. A causa degli affioramenti rocciosi discontinui e della profondità diversa del suolo, i pascoli analizzati si presentano come un mosaico composto dalle varie fasi dinamiche in cui si trova lo strato erbaceo.
Le leguminose, soprattutto Hippocrepis comosa, ed alcune specie come Helianthemum oelandicum e Thymus longicaulis hanno un ruolo determinante nelle prime due fasi di evoluzione, mentre le graminacee assumono il ruolo di dominanti nella terza fase (40% della fitomassa totale).
Si può ipotizzare che la presenza massiccia dei muschi e di uno strato continuo di necromassa condizionino il successo della germinazione dei semi, in quanto ad alti valori di fitomassa briologica e di necromassa (caso della fitocenosi 1) si registra una percentuale maggiore di plantule in quasi tutte le popolazioni. Il maggior sviluppo di muschi e licheni nella fitocenosi 1 si può spiegare, almeno in parte, con la presenza di un microclima favorevole, dovuto al tappeto vegetale alto e chiuso, e dalle differenze di topoclima sulle due esposizioni.
Tra le due fitocenosi comparate sono state messe in evidenza differenze significative per ben 11 componenti della materia organica totale, anche per le specie edificatrici come Sesleria nitida, Festuca trachyphylla, Bromus erectus, Hippocrepis comosa, Anthyllis vulneraria, ecc.
La struttura dei numeri evidenzia che le graminacee sono più numerose nella fitocenosi 1, mentre le leguminose e "altre fanerogame" sono prevalenti nella fitocenosi 2.
Le popolazioni di leguminose e di "altre fanerogame" presentano nella fitocenosi 2 una percentuale di individui fertili superiore a quella degli sterili, mentre le graminacee hanno generalmente pochi individui fertili in ambedue le fitocenosi. La superiorità del rapporto fertili/sterili nella fitocenosi 2 si manifesta anche a livello di singole popolazioni ed è probabilmente determinata dall’elevata concorrenza che invece caratterizza la fitocenosi 1.
Nelle fasi avanzate di evoluzione, la struttura sotteranea delle fitocenosi è ben sviluppata a causa delle grandi radici delle graminacee e per la presenza di rizomi e bulbi che compaiono in grande quantità ed a livelli stratificati, in quanto lo strato di necromassa si alza progressivamente.
Il sistema sotteraneo complesso della fitocenosi 1 avvolge in una rete molto fitta l’intero volume di suolo e numerosi frammenti di roccia, realizzando una buona protezione contro l’erosione e favorendo i processi pedogenetici.
Lo studio di indici popolazionistici quali numero d’individui, frequenza e fitomassa per le 59 specie, evidenzia che circa il 25-50% di queste formano popolazioni con numero di effettivi e fitomassa piuttosto elevati (> 1 ind. m-2 e > 1 g m-2). Esiste un gruppo di specie edificatrici comuni ad ambedue le fitocenosi (Sesleria nitida, Helianthemum oelandicum, Festuca trachyphylla, Thymus longicaulis e Carex macrolepis) con numero di individui, frequenze e fitomassa elevati, e due gruppi di specie differenziali, il primo con alto numero di effettivi ed elevata fitomassa nella fitocenosi 1 (Chamaecytisus hirsutus, Phleum ambiguum e Anthoxanthum odoratum), ed il secondo con valori elevati degli stessi indicatori nella fitocenosi 2 (Hippocrepis comosa, Koeleria splendens e Asperula purpurea).